李氏禾湿地系统如何净化污染水体(多图)
需要咨询购买水处理设备?免费为您提供产品报价和设计方案,咨询电话:010-8022-5898 摘要:利用湿生铬超富集植物李氏禾构建了三段式波形潜流式人工湿地,并以相同设计的无植物湿地系统作为对照,比较研究了李氏禾湿地系统对Cr(Ⅵ)的净化效果.同时,综合运用电子顺磁共振(EPR)、X光电子能谱(XPS)、傅里叶变换红外光谱(FTIR)等手段,并结合有机质含量、pH、Eh和Cr质量平衡分析,探讨了李氏禾湿地系统对Cr(Ⅵ)的去除机理.结果显示,李氏禾湿地系统对Cr(Ⅵ)的去除率显著高于对照湿地,且李氏禾湿地基质中的有机质含量是对照湿地的3倍.这表明李氏禾增加了湿地基质中的有机质含量,从而提高了湿地系统对Cr(Ⅵ)的净化能力.李氏禾组织中的Cr主要以Cr(Ⅲ)形式存在,其中,茎部和叶部未检测到Cr(Ⅵ),而仅有1.95%的Cr以Cr(Ⅵ)形式存在于根部中.EPR分析结果显示,李氏禾叶部中的Cr(Ⅲ)以有机酸结合态的形式存在,根部中的Cr(Ⅲ)以氢氧化物的形式存在,这表明李氏禾能有效地将Cr(Ⅵ)还原成Cr(Ⅲ).基质表面吸附的Cr大部分为Cr(Ⅲ),而Cr(Ⅵ)仅占4.99%.XPS分析进一步证明,基质表面同时吸附了Cr(Ⅲ)和Cr(Ⅵ),但主要以Cr(Ⅲ)为主,且Cr(Ⅲ)可能以氢氧化物的形式存在.由此判断,进入湿地的Cr(Ⅵ)在基质中被还原成Cr(Ⅲ).FTIR分析结果显示,基质表面的羟基、氨基、羧基、C O、C—O、C—H等基团与Cr的吸附有关,这表明湿地基质中的有机质作为电子供体参与了Cr(Ⅵ)的还原.李氏禾湿地系统能将水体中可溶性的Cr(Ⅵ)有效地还原成难溶的Cr(Ⅲ)化合物,并将其转化成湿地生态系统物质循环的惰性部分. 1 引言 铬(Cr)是一种广泛应用的工业原料,电镀、皮革加工等过程都会产生含Cr污水进而造成水体污染.水体中的Cr主要以Cr(Ⅵ)和Cr(Ⅲ)两种稳定的价态存在,而Cr(Ⅵ)和Cr(Ⅲ)在毒性、移动性和生物有效性上存在着明显的差异.Cr(Ⅵ)通常以CrO2-4和Cr2O2-7形式存在,有毒且可溶,被认为是致畸和致癌物质.由于CrO2-4、Cr2O2-7带负电(pH>3),不易被矿物质和有机物质吸附,因而Cr(Ⅵ)在环境中移动性很高.相反,Cr(Ⅲ)难溶解且不易移动,毒性为Cr(Ⅵ)的1/100.因此,将Cr(Ⅵ)还原成Cr(Ⅲ)是减少水体Cr污染的重要途径. 目前,清除水中Cr(Ⅵ)污染的方法有电解还原法、化学沉淀法、离子交换法和微生物还原法等,但这些方法需要消耗大量的能源和试剂且成本较高,不适于大面积、低浓度的Cr(Ⅵ)污染水体处理.人工湿地相对上述方法具有低成本、低能耗和生态服务功能等优点,因此,被认为是一种经济有效的Cr(Ⅵ)去除途径. 李氏禾(Leersia hex and ra Swartz)是中国境内首次发现的湿生铬超富集植物,对Cr(Ⅵ)和Cr(Ⅲ)有很强的耐受和富集能力,并且具有生长迅速、根系发达、易于人工种植等优点.因此,该植物在水体Cr(Ⅵ)污染修复中极具应用前景.在前期实验中,我们发现以李氏禾作为湿地植物构建的三段式波形潜流式人工湿地对Cr(Ⅵ)污染水体具有很强的净化能力.然而,该湿地系统对水体Cr(Ⅵ)的去除机理仍不十分清楚. 因此,本研究以实验室规模的李氏禾湿地系统为对象,综合运用电子顺磁共振(EPR)、X光电子能谱(XPS)、傅里叶变换红外光谱(FTIR)等手段,对湿地出水水质、铬的价态变化及湿地基质和植物中铬的化学特征进行分析,探讨李氏禾湿地系统净化水体Cr(Ⅵ)的生物地球化学机理,以期为提高湿地的净化性能提供科学依据. 2 材料与方法 2.1 装置与方法 实验于温室大棚中进行,采用以PVC板材粘合成的三段式波形潜流式人工湿地(1.3 m×0.5 m×0.3 m,长×宽×高,图 1).其中,进水区长0.1 m,湿地长1.2 m,湿地平分为3段,每段长0.4 m.设计的表面负荷率为0.1 m3 · m-2 · d-1,进水流量为0.06 m3 · d-1,水力停留时间为1 d.进水区填充高0.28 m的砾石(粒径2 cm左右),基质深度为0.25 m,其中,下层为0.05 m的砾石(粒径1 cm左右),上层为0.2 m的稻田土+泥炭混合基质(稻田土 : 泥炭=2 : 1,体积比,粒径0.9 mm左右).湿地植物——李氏禾从桂林市郊采集,以80株 · m-2成行种植于基质表面.李氏禾中的铬含量未检测到,基质中的铬含量为(5.47±0.15)mg · kg-1,加入到湿地中的基质质量约为132.0 kg,基质中的有机质含量为3.50%±0.06%,pH为7.52±0.02,Eh为(392.7±4.5)mV.
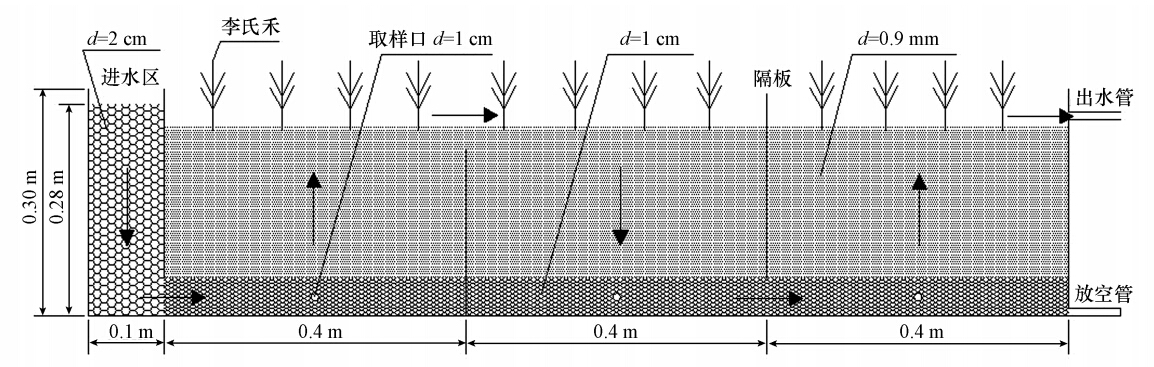
图 1 三段式波形潜流式人工湿地示意图 2012年4~6月为李氏禾生长期,李氏禾生长至6月末基本布满整个湿地.湿地从2012年7月开始运行,Cr(Ⅵ)污染水体由K2Cr2O7配制而成,进水中Cr(Ⅵ)浓度更换梯度依次为2.50、3.75、5.00、7.50、10.0 mg · L-1,采用连续进水的方式,运行至2012年12月.实验采用相同设计的无植物人工湿地作为对照,与种植了李氏禾的湿地进行对比研究. 2.2 取样方法及测试项目 湿地运行结束后,采用梅花形布点法采集两组湿地的基质及李氏禾植株的根、茎、叶.将根、茎、叶洗净后一部分置于105 ℃烘箱中杀青30 min,然后80 ℃烘干至恒重,粉碎后用于Cr的测定;另一部分充分剪碎后置于研钵中,加入液氮,迅速研磨成粉末状,用于EPR的检测.将土样置于105 ℃烘箱中烘干至恒重,分别研磨过筛后,李氏禾湿地的基质一部分用于有机质含量、pH、Eh和Cr的测定,另一部分用于XPS、FTIR的检测;对照湿地的基质测定其有机质含量、pH和Eh.其中,根、茎、叶和基质中Cr(Ⅵ)的提取采用碱式消解法(EPA3060A),将萃取后得到的Cr(Ⅵ)用二苯碳酰二肼分光光度法进行测定;总Cr的提取采用酸式消解法,将消解后得到的Cr用火焰原子吸收分光光度法进行测定;有机质含量的测定采用重铬酸钾容量法;pH、Eh的测定采用电极法.测量实验均设置3个平行样,且取其数据的平均值±标准差作为实验结果. 2.3 实验主要仪器 实验仪器主要包括:Sartorius PB-10型pH计、ORP复合电级、TRF-2系列土壤测试仪、SHZ-B型水浴恒温振荡器、UV-9600紫外可见分光光度计、AA-700原子吸收分光光度计、JES-FA200电子顺磁共振波谱仪、Magna-IR750傅里叶变换红外光谱仪、Axis Ultra多功能成像电子能谱仪. 3 结果 3.1 李氏禾湿地系统对Cr(Ⅵ)的去除率及基质的有机质含量、pH和Eh 从表 1可以看出,对照湿地基质中的有机质含量从3.50%降低至1.12%,说明湿地系统在去除Cr(Ⅵ)的过程中有机质在逐渐被消耗.而李氏禾湿地基质中的有机质含量依然能维持在较高水平(3.36%),是对照湿地的3倍,且在Cr(Ⅵ)进水浓度为10 mg · L-1时,李氏禾湿地系统对Cr(Ⅵ)的去除率比对照湿地高出11.39%.这说明李氏禾增加了湿地基质中的有机质含量,从而提高了湿地系统对Cr(Ⅵ)的净化能力.两组湿地基质的pH均接近中性(表 1),结合表 1中的Eh可知,两组湿地基质中的Cr均主要以Cr(Ⅲ)形式存在. 表1 两组湿地系统对Cr(Ⅵ)的去除率及基质的有机质含量、pH和Eh
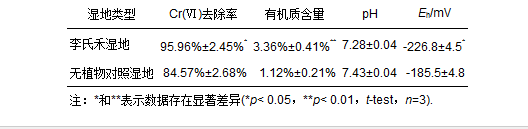
3.2 李氏禾湿地系统铬质量平衡 从图 2可以看出,从李氏禾湿地系统中流出的铬仅为进入到湿地系统中铬的1.97%,说明进入到湿地系统中铬绝大部分被截留在了湿地系统中.截留在基质中的铬占到了进入湿地系统中铬的83.88%,李氏禾中的铬仅占到14.15%,说明截留在湿地系统中的铬绝大部分存在于基质中.基质中的Cr(Ⅵ)仅占4.99%,Cr(Ⅲ)占到了78.89%,说明基质中的Cr大部分为Cr(Ⅲ).李氏禾的茎部和叶部未检测到Cr(Ⅵ),Cr(Ⅲ)占到了1.80%;李氏禾根部中的Cr(Ⅵ)仅占1.95%,Cr(Ⅲ)占到了10.40%,可知李氏禾植株中的Cr(Ⅵ)仅占1.95%,Cr(Ⅲ)占到了12.20%.这说明李氏禾植株中的Cr主要以Cr(Ⅲ)形式存在.
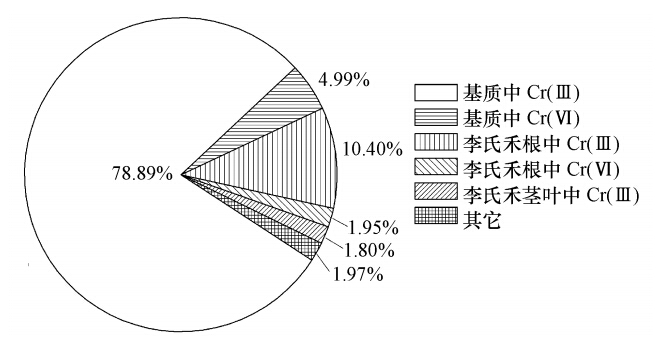
图 2 铬在李氏禾湿地系统各介质中的分布 3.3 李氏禾吸附Cr(Ⅵ)后EPR谱图分析 g为波普分裂因子,能反映出一种物质分子内局部磁场的特征,与过渡金属离子的d电子壳层充满程度有关,且不同价态Cr的d电子壳层充满程度有所不同,其可以确定是否有同一种物质存在.由图 3可知,标准物质K2Cr2O7的EPR谱图在整场磁场范围内没有出现明显特征峰.标准物质CrCl3和Cr(OH)3的EPR谱图在g=2.00处均有明显的信号区域.标准物质C6Cr2O12 · 6H2O的EPR谱图在g=2.20处有明显的信号区域且同时在g=4.00处出现特征峰. 李氏禾叶部的EPR谱图在g=4.00处出现特征峰,且与C6Cr2O12 · 6H2O特征峰g值一致.由于有机铬化物如草酸氢铬、柠檬酸铬、Cr(Ⅲ)-EDTA在g=4.00~5.50均出现特征峰,因此,李氏禾叶部中的Cr(Ⅲ)以有机酸结合态的形式存在.在g=2.00处附近出现一系列超精细谱线可以归结于Mn(Ⅱ)和C,但也不排除此信号区域还存在少量的Cr(Ⅲ)信号,因为其出现信号区域的g值与Cr(OH)3产生信号区域的g值一致,所以推测还可能存在Cr(OH)3.李氏禾根部的EPR谱图在g=2.00处出现明显的信号区域,与标准物质Cr(OH)3产生明显信号区域的g值一致,说明李氏禾根部中的Cr(Ⅲ)以氢氧化物的形式存在.结合李氏禾植株中Cr(Ⅵ)仅占1.95%、Cr(Ⅲ)占到了12.20%(图 2)可知,李氏禾能有效地将Cr(Ⅵ)还原成Cr(Ⅲ).此外,李氏禾根部EPR谱图在g=2.00和g=1.98处出现两个连续的信号区域,峰宽为3.89 mT.研究表明,Cr(Ⅴ)顺磁信号存在的位置于g≈2.00周围且峰宽<6 mT,说明李氏禾根部中还同时存在少量的中间价态Cr(Ⅴ).
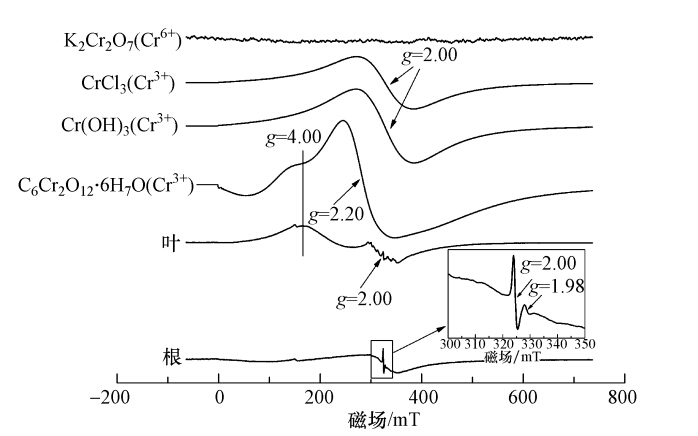
图 3 李氏禾叶部和根部EPR谱图 3.4 李氏禾湿地基质吸附Cr(Ⅵ)前后的表面分析 3.4.1 XPS分析 XPS宽扫描谱图分析:XPS宽扫描可以给出除H和He以外所有元素的内层电子的结合能,通过元素的特征结合能与灵敏度因子可以确定表面的化学成分及其相对含量.由图 4a可知,基质在531.88 eV处出现了强峰,表明其主要组成为O;在284.63 eV和102.83 eV处出现弱峰,表明其主要组成为C和Si.此外,图 4a中Al、Ca、Fe元素的峰可辨.由图 4b可知,基质吸附Cr(Ⅵ)后在577.87 eV处出现了Cr2p3/2特征峰,说明Cr吸附在湿地基质表面.截留在基质中的铬占到了进入湿地系统中铬的83.88%(图 2),表明进入湿地的Cr(Ⅵ)主要通过基质的吸附而得到去除.
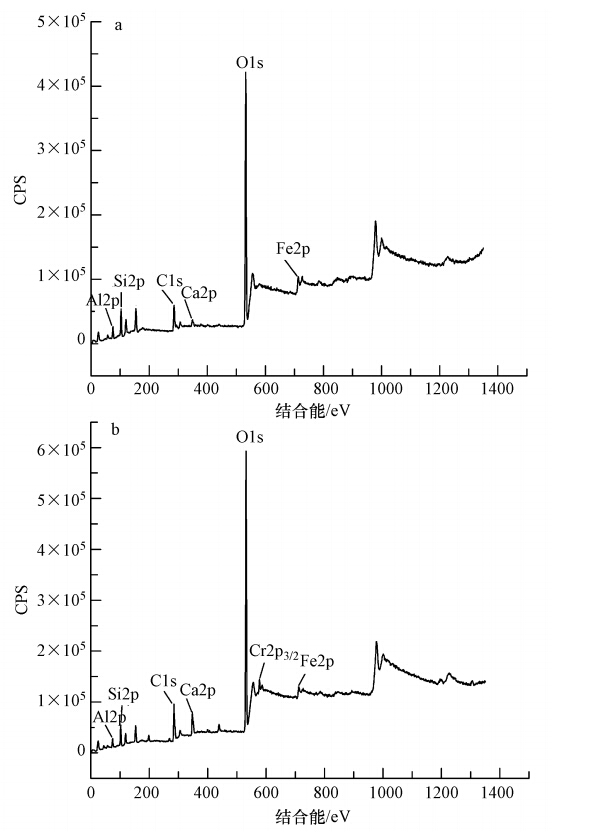
图 4 李氏禾湿地基质吸附Cr(Ⅵ)前(a)、后(b)宽扫描XPS谱图 Cr2p高分辨率XPS谱图分析:过渡金属元素常常可以通过主要分裂线(2p1/2和2p3/2,3d3/2和3d5/2等)的能级间距、能量位置和谱线的形状来进行化学价态的鉴定.由图 5可知,在577.87 eV和587.12 eV处出现很明显的特征峰,分别代表Cr2p3/2和Cr2p1/2轨道的电子结合能.对Cr2p3/2轨道的特征峰进行分峰,得到结合能分别处在577.29 eV和579.72 eV处的两个峰,其中,579.72 eV处为一较小肩峰.因为Cr(Ⅲ)的2p3/2轨道结合能通常在576.8~577.7 eV之间,一般不大于578 eV;Cr(Ⅵ)的2p3/2轨道结合能通常在579.0~581.0 eV之间,一般大于578 eV(Park et al., 2007).因此,图 5中577.29 eV处的特征峰为Cr(Ⅲ)2p3/2轨道结合能对应的特征峰,579.72 eV处的特征峰为Cr(Ⅵ)2p3/2轨道结合能对应的特征峰.由于XPS峰面积能客观反应出物质表面元素价态的相对含量,因此,从图 5可以看出,基质表面Cr(Ⅵ)含量相对很低,Cr主要为Cr(Ⅲ),与图 2基质中Cr(Ⅵ)仅占4.99%、Cr(Ⅲ)占到了78.89%的结果相一致,表明进入湿地的Cr(Ⅵ)在基质被还原成Cr(Ⅲ).研究表明,Cr(OH)3中Cr—O键的Cr2p3/2峰位于577.3 eV处,与湿地基质表面Cr(Ⅲ)2p3/2特征峰的轨道结合能非常接近.这表明基质表面Cr(Ⅲ)的结合状态可能为Cr(OH)3中Cr的结合状态,即Cr(Ⅲ)可能以氢氧化物形式存在.同时,基质吸附Cr(Ⅵ)前后均存在Fe元素(图 4),推测Fe(Ⅱ)对Cr(Ⅵ)的还原可能起到了一定的作用.
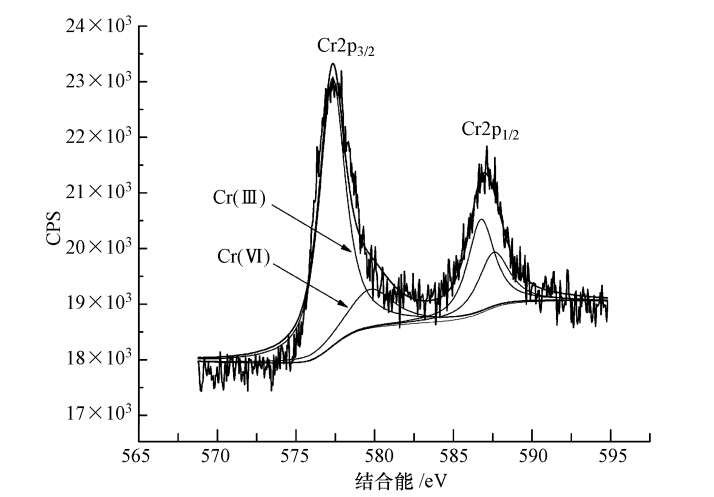
图 5 李氏禾湿地基质吸附Cr(Ⅵ)后Cr2p高分辨率XPS谱图 3.4.2 FTIR分析 由图 6可知,湿地基质吸附Cr(Ⅵ)之前在3430 cm-1处存在一个强而宽的谱带,说明基质表面存在O—H、N—H伸缩振动;2930 cm-1处的吸收峰与—CH2的反对称伸缩振动相关;2360 cm-1处为空气中CO2吸收峰;1630 cm-1处的吸收峰与C O伸缩振动相关;1420 cm-1处的吸收峰与羧基中的C O伸缩振动相关;1260 cm-1处的吸收峰与C—O伸缩振动相关;1030 cm-1处有强吸收峰,可能表现为Si—O—Si的对称伸缩振动;876、795、692 cm-1处的吸收峰与C—H的弯曲振动相关.基质吸附Cr(Ⅵ)后,3430 cm-1处的吸收峰向高波数区移动了10 cm-1,说明羟基和氨基可能参与了Cr的吸附;1420 cm-1处的吸收峰向高波数区移动了30 cm-1并形成强吸收带,说明羧基可能与Cr(Ⅲ)发生了配位作用;1630 cm-1处的吸收峰向低波数区移动了80 cm-1并形成强吸收带,同时1260 cm-1处的吸收峰消失,692 cm-1处的吸收峰向低波数区移动了17 cm-1,表明C O、C—O、C—H等基团与Cr的吸附有关.这些结果表明,湿地基质中的有机质作为电子供体参与了Cr(Ⅵ)的还原.
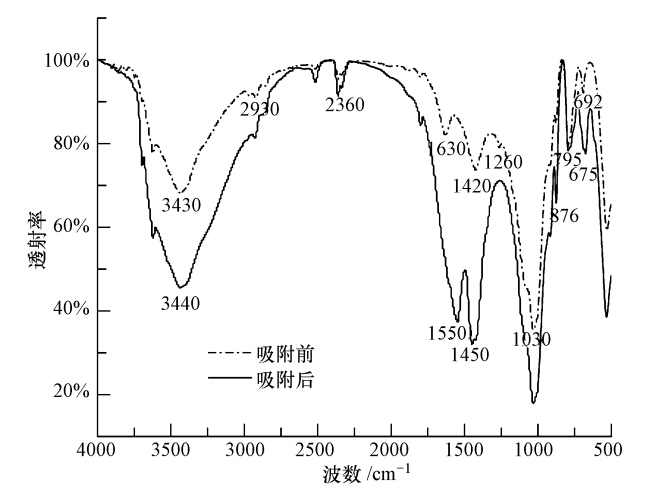
图 6 李氏禾湿地基质吸附Cr(Ⅵ)前后FTIR谱图 4 讨论 李氏禾在湿地系统净化Cr(Ⅵ)污染水体过程中的贡献主要体现在增加基质中的有机质含量,从而提高湿地系统的净化能力(表 1).因为植物的凋落物、根际分泌物及腐烂的根经矿化后都会增加基质中的有机质含量.其具体包括:①植物的凋落物和腐烂的根经矿化后会增加基质中腐殖酸和富里酸的含量;②根际分泌物的代谢活动会产生乳酸、扁桃酸、酒石酸、草酸、水杨酸等有机酸.除此之外,李氏禾在湿地系统净化Cr(Ⅵ)污染水体过程中的贡献还体现在能吸收一定量的铬(图 2),并能有效地将Cr(Ⅵ)还原成Cr(Ⅲ)(图 3).其中,叶部中的Cr(Ⅲ)以有机酸结合态的形式存在,与Howe等的研究结果:短萼车轴草叶部中的Cr(Ⅲ)以有机酸结合态的形式存在相一致.这可能是因为Cr(Ⅲ)以有机酸络合物的形式螯合成毒性较低的稳定物质储存于细胞非功能区,从而减少游离的Cr(Ⅲ)对李氏禾叶部的毒害作用.根部中的Cr(Ⅲ)以氢氧化物的形式存在,可能是由于进入根部的Cr(Ⅵ)转化成Cr(Ⅲ)后,被根部细胞表面的果胶和细胞质外体中的负电位基团吸收,进而形成Cr(OH)3;也有可能是Cr(Ⅲ)进入根部细胞质(pH=7.3~7.6)后发生了沉淀.另外,根部还存在少量的中间价态Cr(Ⅴ),是因为进入根部的Cr(Ⅵ)还原成Cr(Ⅲ)时会先经过中间价态Cr(Ⅴ). 湿地基质在湿地系统净化Cr(Ⅵ)污染水体过程中起主要作用,不仅能吸附大部分的Cr(Ⅵ)(图 2),而且还能有效地将Cr(Ⅵ)还原成Cr(Ⅲ)(图 5),其中,有机质作为电子供体直接参与了Cr(Ⅵ)的还原(图 6).研究分发现,根际分泌物和根际微生物的代谢活动产生的乳酸、扁桃酸、酒石酸、草酸、水杨酸等有机酸能处进Cr(Ⅵ)的还原,土壤里的腐殖酸和富里酸也能对Cr(Ⅵ)进行还原和固定,而这些还原固定Cr(Ⅵ)的过程主要是通过有机质所含有的类似羧基和酚羟基等官能团与金属离子发生配位作用的方式来实现的.这与本实验中湿地基质表面的羧基与Cr(Ⅲ)发生了配位作用,羟基、氨基、C O、C—O、C—H等基团参与了铬的吸附的研究结果相一致.此外,有机质还可能通过间接作用提高了湿地系统对Cr(Ⅵ)的净化能力.主要包括:①根际分泌物产生的有机酸提高了其周围微生物的活性,从而促进了微生物对Cr(Ⅵ)的转化和固定;②植物为基质中的微生物提供了有机质,微生物在消耗有机质时会消耗氧气从而有利于Cr(Ⅵ)的还原和沉降;③在厌氧环境下,硫酸盐还原菌消耗有机质时会产生必要的硫化物,进而与Cr(Ⅵ)发生反应,将Cr(Ⅵ)还原成Cr(Ⅲ)以形成溶解性极小的金属硫化物.不仅有机质参与了湿地基质对Cr(Ⅵ)的还原,而且湿地基质中的矿物质可能也参与了Cr(Ⅵ)的还原.本实验研究中Fe(Ⅱ)对Cr(Ⅵ)的还原可能起到了一定的作用,其还原过程推测是Fe(Ⅱ)将水体中的Cr(Ⅵ)还原成了Cr(Ⅲ),进而在中性条件下以Cr(OH)3及铁铬氧化水合物[FexCr1-x(OH)3]的形式存在于基质中.这样溶解性较低的Cr(Ⅲ)被紧紧地吸附在基质表面,导致铬从液相进入到固相,从而减少了水体中Cr(Ⅵ)浓度.具体参见污水宝商城资料或http://www.dowater.com更多相关技术文档。 5 结论 李氏禾增加了湿地基质中的有机质含量,从而提高了湿地系统对Cr(Ⅵ)的净化能力.李氏禾组织中的Cr主要以Cr(Ⅲ)形式存在,其中,茎部和叶部未检测到Cr(Ⅵ),而仅有1.95%的铬以Cr(Ⅵ)形式存在于根部中;叶部中的Cr(Ⅲ)以有机酸结合态的形式存在,根部中的Cr(Ⅲ)以氢氧化物的形式存在.基质表面同时吸附了Cr(Ⅲ)和Cr(Ⅵ),Cr(Ⅵ)仅占4.99%,大部分为Cr(Ⅲ),且Cr(Ⅲ)可能以氢氧化物的形式存在.有机质作为电子供体参与了湿地基质对Cr(Ⅵ)的还原,这表明李氏禾湿地系统中的植物和基质将水体中可溶性的Cr(Ⅵ)阴离子还原成溶解性较低的Cr(Ⅲ)阳离子,并吸附在基质表面的羟基、氨基、羧基、C O、C—H、C—O等基团上,从而使水体中的Cr(Ⅵ)浓度下降. (责任编辑:李德馨) |